腺苷(ADO,adenosine)是一种无所不在、代谢迅速的嘌呤核苷,生理半衰期仅为几秒钟。因此,体内循环ADO的浓度很难测量。细胞外ADO(eADO)的生理浓度在纳摩尔水平,但在病理条件下,它们可能高达100mM。鉴于ADO的不稳定性,它主要通过自分泌和旁分泌信号发挥作用。它参与细胞能量转移,因为它是形成二磷酸腺苷(ADP)和三磷酸腺苷(ATP)的基础。此外,它在各种信号转导途径中发挥重要作用,是形成环磷酸腺苷(cAMP)等信号分子的一个组成部分。

细胞外ADO的主要来源是ATP分子,ATP被几种细胞类型的一系列膜定位酶水解为ADO,尤其是胞外核苷酸三磷酸二磷酸水解酶1(CD39)、胞外-5′-核苷酸酶(CD73)、外核焦磷酸酶/磷酸二酯酶(ENPP)和前列腺酸性磷酸酶(PAP)。eADO的产生也可能通过内在代谢途径进行,主要涉及腺苷激酶(ADK)、S-腺苷同型半胱氨酸水解酶(SAHH)、细胞质5′-核苷酸酶-I(cN-I)和细胞表面环ADP核糖水解酶(CD38)的NAD+补救途径。
从最初的炎症/局部组织损伤到癌前生态位和已发展的肿瘤,腺苷过量产生发生在肿瘤发生的所有阶段,使腺苷能途径成为一个有吸引力但具有挑战性的治疗靶点。目前,免疫肿瘤学领域的许多研究都集中在恢复免疫监测上,主要是通过单药或联合用药阻断肿瘤微环境(TME)中的腺苷产生酶和免疫细胞上的腺苷受体。
来自细胞外基质(ECM)的信号除了在细胞间通讯中的作用外,还相互影响健康的组织结构和相关的组织特异性功能,这个过程被称为双向组织微环境动态相互作用。在癌前生态位发生初始转化后,随着突变的积累,分子异质性增加,这就造成了代谢相互作用的异质性。TME内所有类型的细胞都面临着一些挑战,包括间质压力升高、对氧气和营养素的需求增加、供应输送受损以及代谢产物清除效率低下。反过来,TME内细胞代谢的变化可能以支持肿瘤维持和加速TME重塑的方式指导与ECM的相互作用。
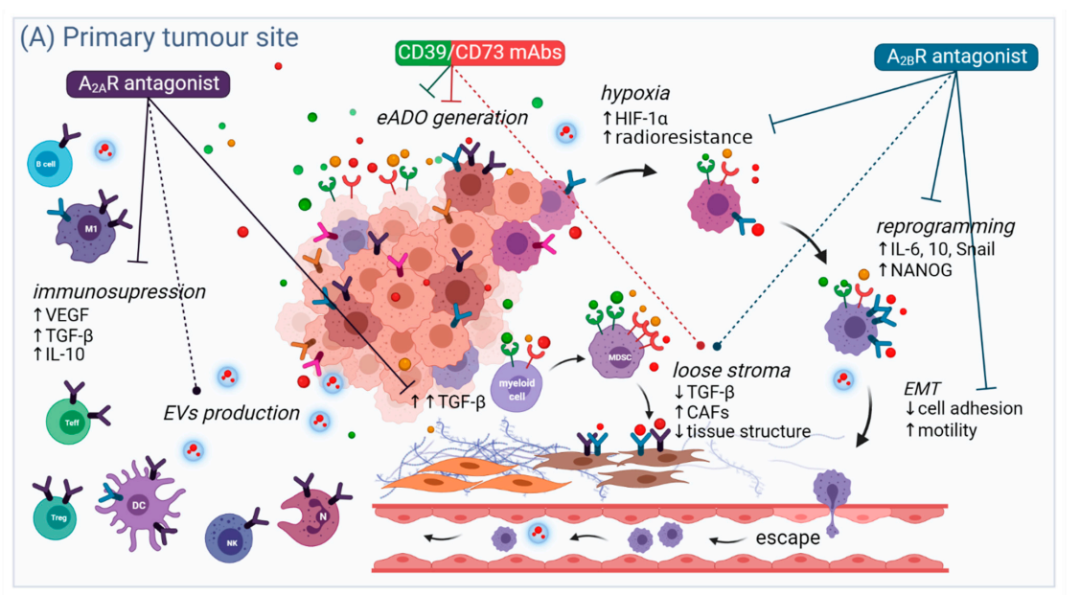
嘌呤释放到细胞外环境在细胞间通讯中起着重要作用。在TME中,组织损伤或化疗导致的死亡癌细胞的免疫原性通过ATP水解为ADO而被中和。此外,不仅癌细胞释放eADO以维持特定的免疫抑制表型,事实上,TME内Treg细胞提供ATP分子和CD39/CD73胞外酶来维持富含ADO的TME,进而通过A2AR触发相邻效应细胞的免疫抑制。
在浸润性中性粒细胞中观察到另一个有趣的正反馈回路,通过缺氧诱导因子1α(HIF-1α)依赖机制抑制ENT表达,确保细胞外空间保持高水平的ADO。此外,TME中存在大量癌症相关成纤维细胞(CAF),在结直肠癌中,CAF通过A2BR刺激驱动CD73的高表达,维持TME中ADO浓度的升高。这些结果表明,在癌症中腺苷途径被TME劫持,TME中的所有细胞类型(癌症、基质细胞、内皮细胞和免疫细胞)都通过AR依赖和独立途径受ADO影响。因此,在生理条件下用于减轻免疫系统过度反应和防止组织损伤的ADO最终成为支持无限制肿瘤生长的成分。
针对腺苷途径成分的临床试验数量有所增加,但大多数试验仍处于开发的早期阶段。目前包括诺华、AZ、礼来、BMS、罗氏等在内的各大药企均积极布局着腺苷途径的药物研发,尚处于临床阶段,没有获批上市的药物。
由于腺苷信号途径的复杂性,靶向腺苷疗法存在一些潜在的局限性和挑战。AR属于A类GPCR,主要由与细胞外结合位点结合的配体调节。AR信号的复杂性带来了一些限制,必须克服这些限制才能开发出有效的治疗方法。此外,AR认知的一个基本障碍是缺乏A2BR和A3R晶体结构,无法加深我们对A2BR和A3R构象动力学的理解。
ARs通常由小分子进行药理学调节,多靶点的参与是小分子存在的一个缺点。例如,选择性A2BR激动剂BAY 60-6583最近被证明与另一个靶分子结合,独立于A2BR上调CAR-T细胞活性。因此,如果不仔细评估,就不能排除ADO类似物可能具有多个目标的可能性。